


Miguel A. Reina、Xavier Sala-Blanch、FabiolaMachés、RiánsaresArriazu、Alberto Prats-Galino
はじめに
末梢神経の微細構造のいくつかの特徴をよりよく理解することで、麻酔の臨床診療に役立つ重要な情報が得られる可能性があります。 この章では、末梢神経の結合組織の超微細構造をレビューして、神経周膜拡散バリアとしての役割と局所麻酔への影響の理解を促進します。
筋肉束
神経とその主要な枝(図1 〜へ 3)神経線維(神経束、神経束)の平行な束で構成されています。 束のサイズ、数、およびパターンは、神経間およびそれらの起源からの異なる距離で異なります。 末梢神経の結合組織が除去されると、通常、20個以上の管状構造または束が見られます。
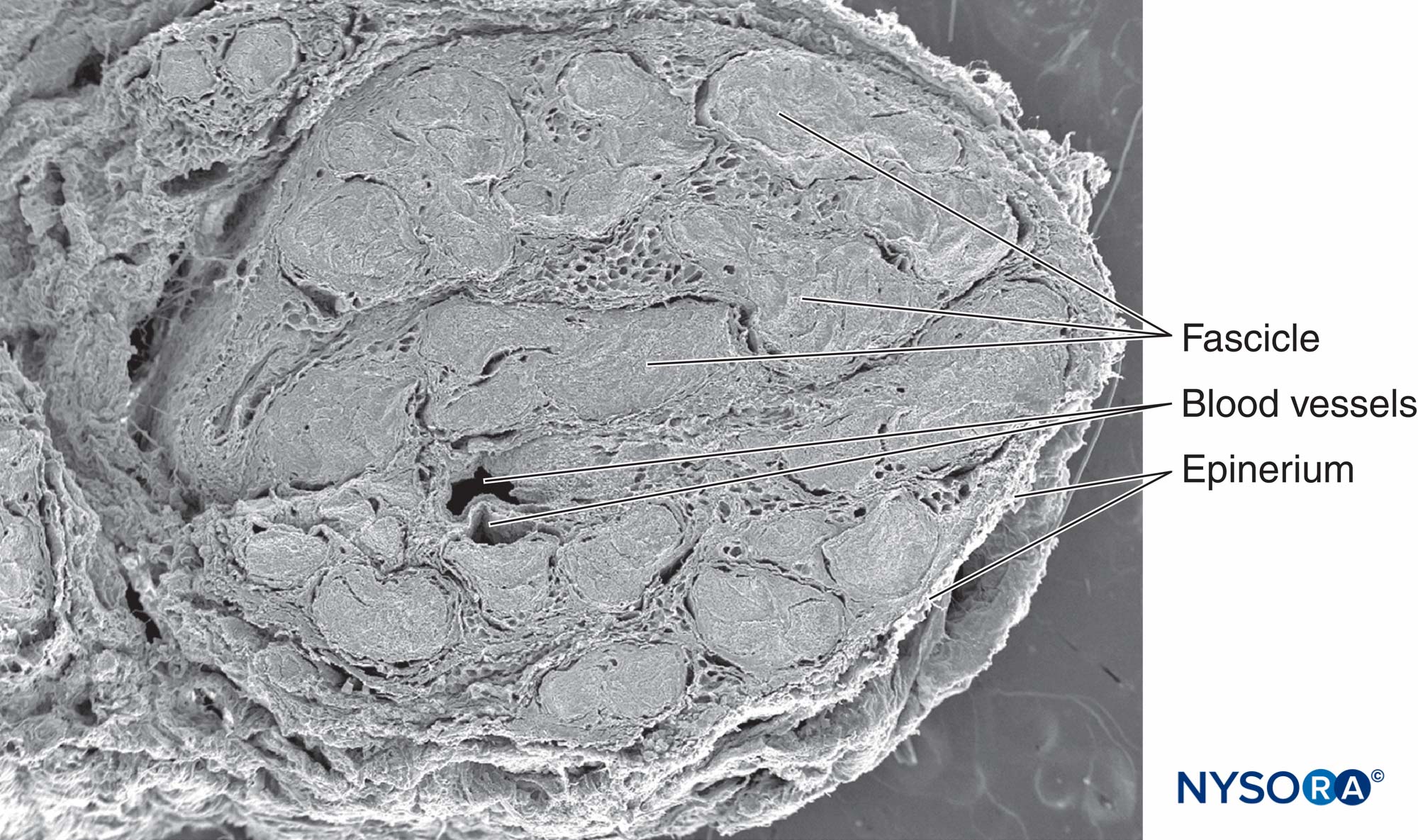
図1 膝窩のレベルでの坐骨神経。 電子顕微鏡で観る。 倍率×25。 (Reina MA、Arriazu R、Collier CBらの許可を得て複製:神経ブロックの実践に臨床的に関連するヒト末梢神経の電子顕微鏡検査。
元の実験および実験データに基づく構造および超微細構造のレビュー、Rev EspAnestesiolReanim。 2013年60月;10(552):562-XNUMX。)
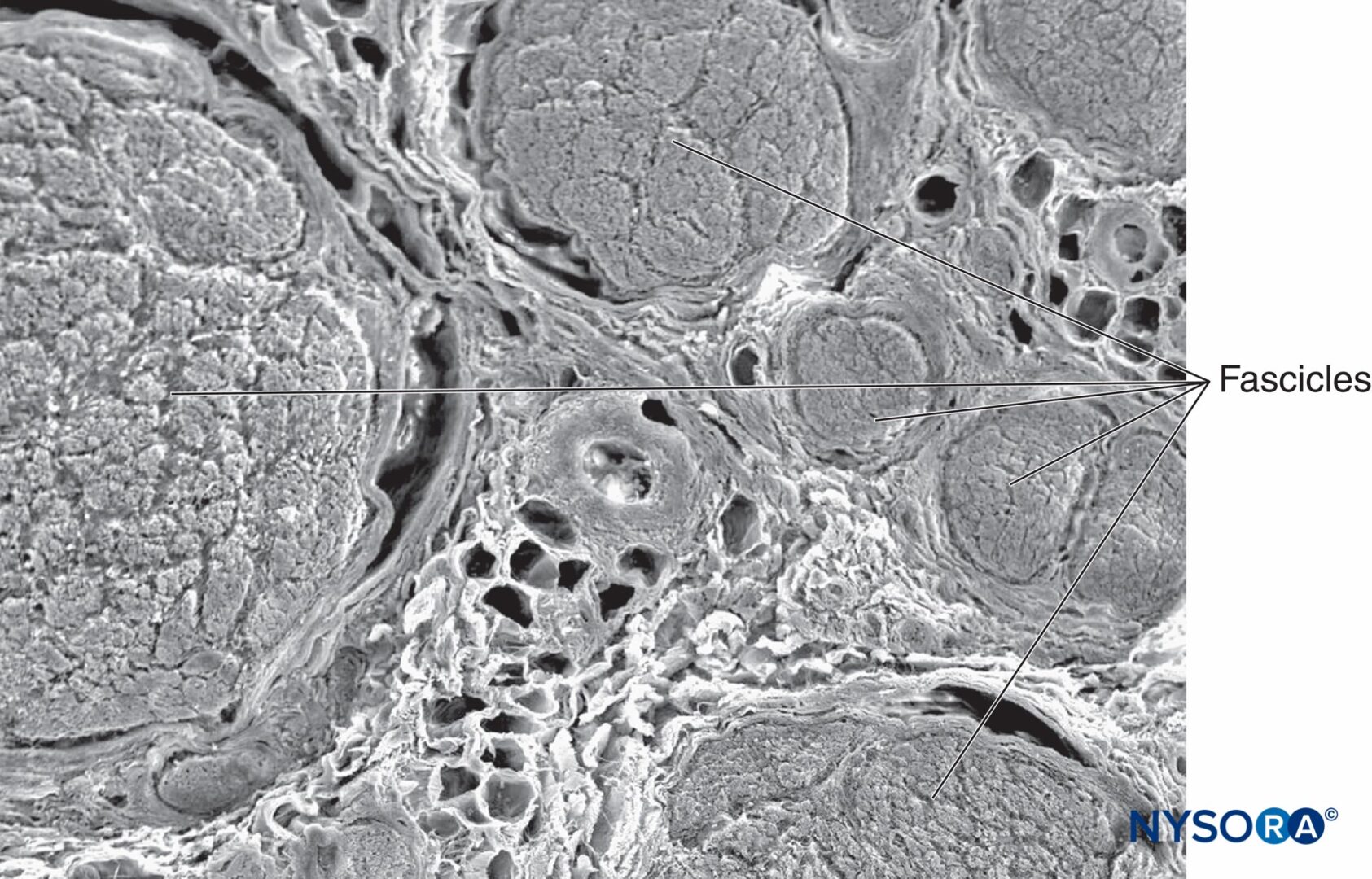
図2 人間の脛骨神経束と束の間の脂肪組織の走査型電子顕微鏡画像。
倍率×75。 (Wikinski J、Reina MA、Bollini Cらの許可を得て複製:Diagnóstico、prevencióny tratamiento delascomplicacionesneurológicasasociadasconlaanestesiaregionalperiféricaycentral。ブエノスアイレス:Panamericana Ed; 2011.)
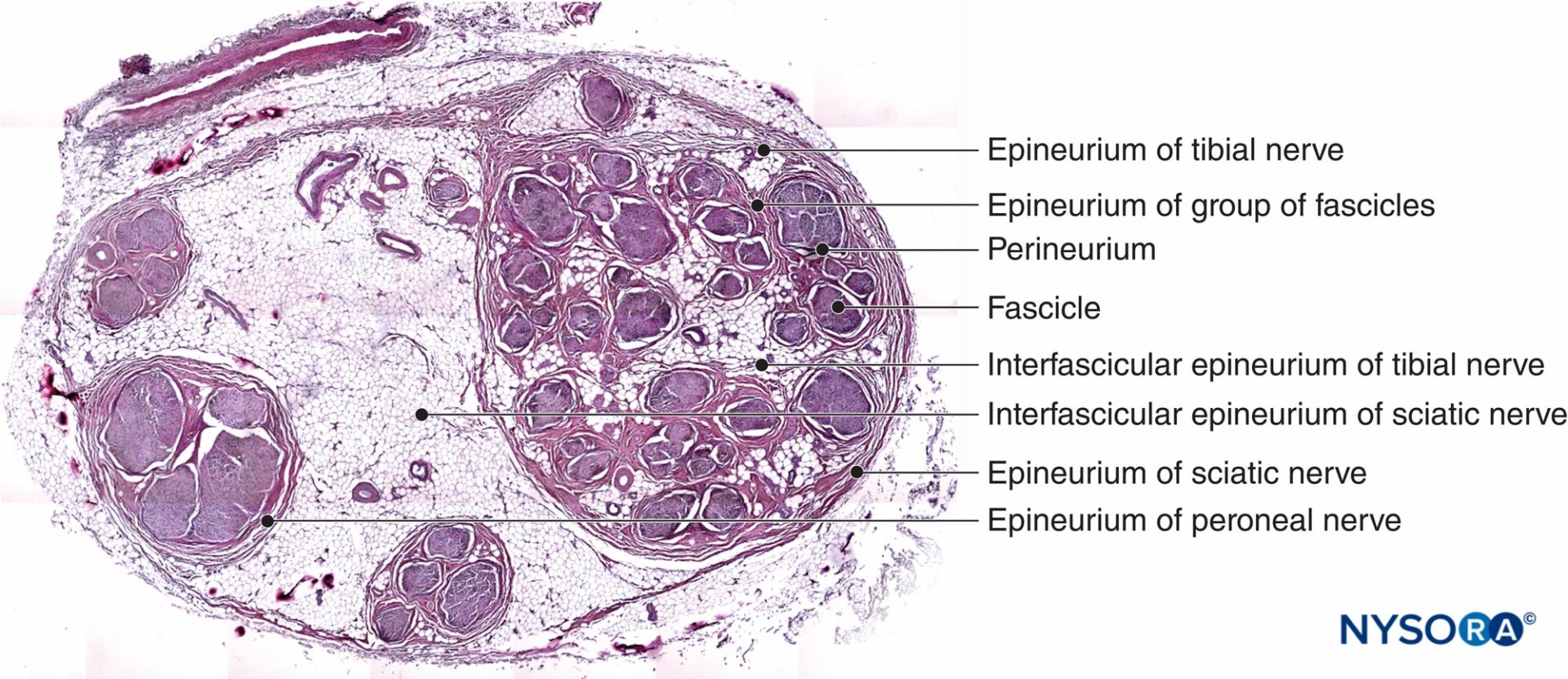
図3 膝窩のレベルでの坐骨神経。 ヘマトキシリン-エオシン。 (Reina MA、De Andres JA、HernándezJMらの許可を得て複製:くも膜下神経根から末梢神経への神経外構造の連続的変化、麻酔ブロックへの影響、および急性術後疼痛の治療。EurJPain。Suppl2011 ; 5(2):377-385。)
各神経の内部で、軸索は、XNUMXつの軸索が神経の長さに沿って異なる束に寄与することができるような方法で神経内神経叢を形成します(図4)。 言い換えれば、軸索は、周辺位置からより中心位置に移動するだけでなく、その下降に沿ってより周辺的に束を完全に交換することができます。 確かに、互いに近い距離にある神経の断面解剖学は、神経内の束の位置と数が非常に変化しやすいことを示しています(参照 図3)神経叢内の存在(図5 & 6)。 末梢神経の束の数、サイズ、および位置も、単一の神経内でも可変であり、神経の長さ4〜5cmに沿って何回も変化する可能性があります。
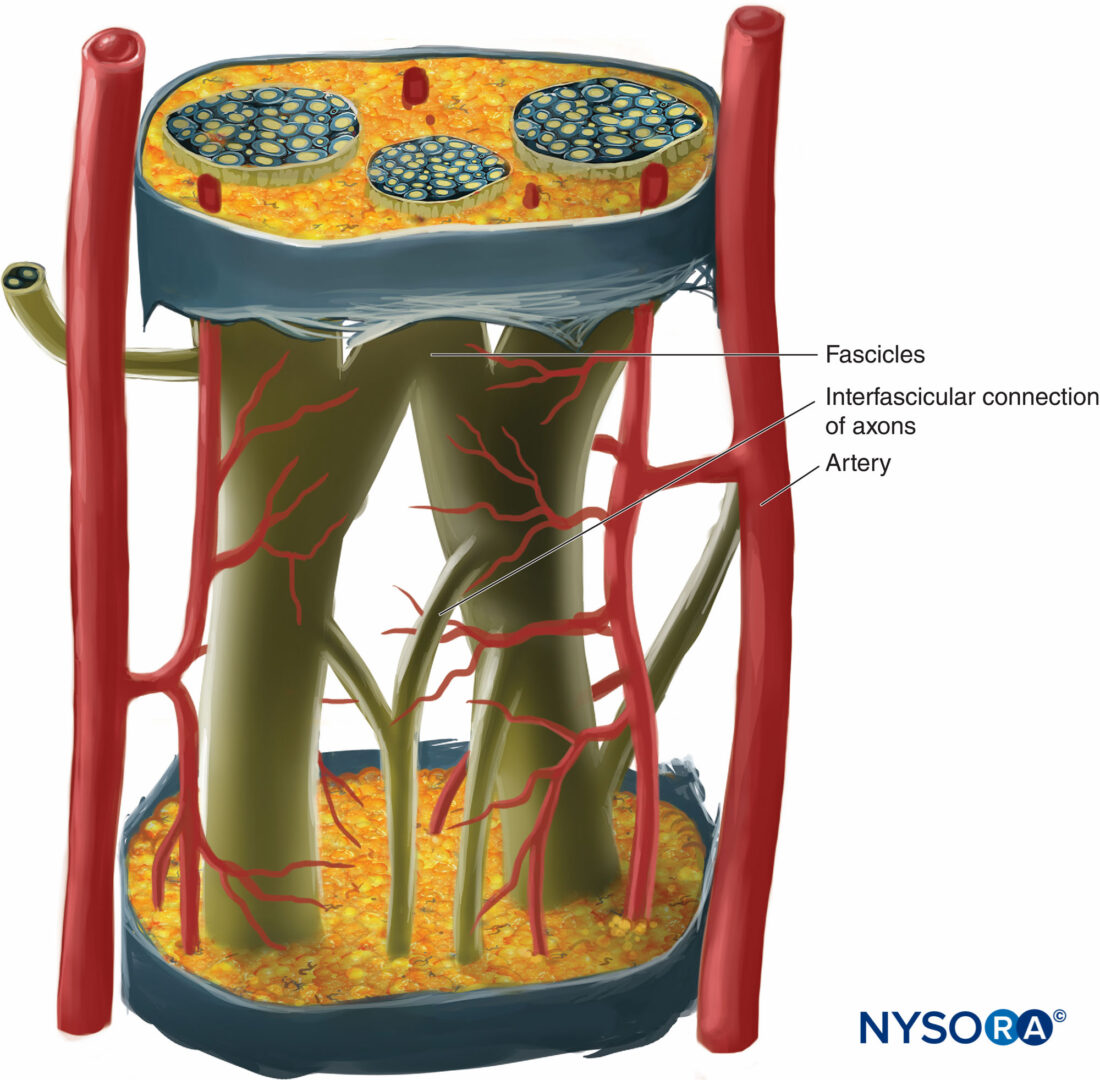
図4。 末梢神経に囲まれた神経内神経叢の図。 (DeAndrésJA、Reina MAの許可を得て複製、
LópezA、et al:Blocsneuruxpériphériques、paresthésieset Injections intraneurales、LePracticieneanAnesthésieRéanimation2010;14:213-221。)
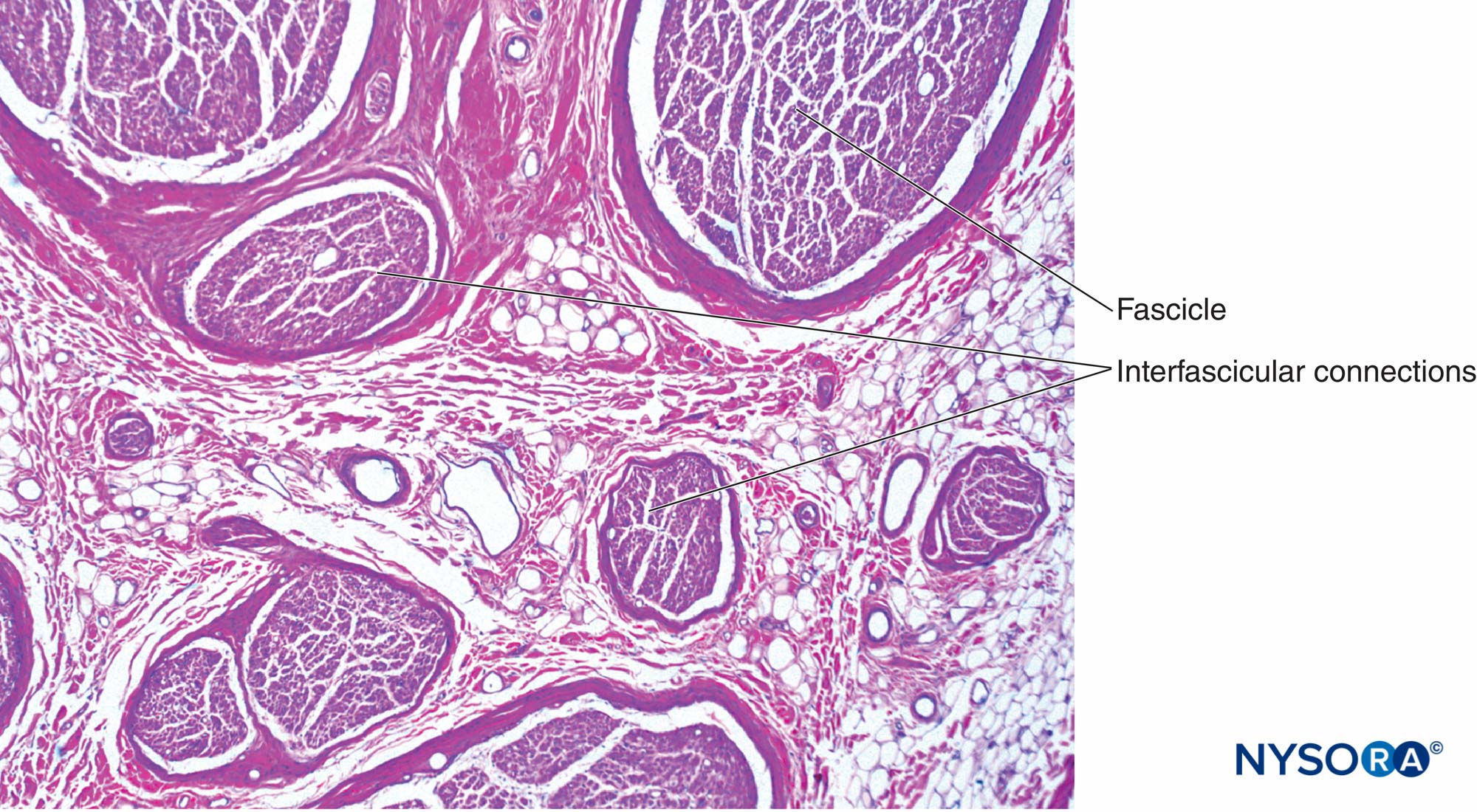
図5 腕神経叢からの末梢神経内の神経内神経叢。 ヘマトキシリン-エオシン。 (Reina MA、Arriazu R、Collier CBらの許可を得て複製:神経ブロックの実践に臨床的に関連するヒト末梢神経の電子顕微鏡検査。元の実験および実験データに基づく構造的および超構造的レビュー、Rev Esp Anestesiol Reanim 。2013年60月;10(552):562-XNUMX。)
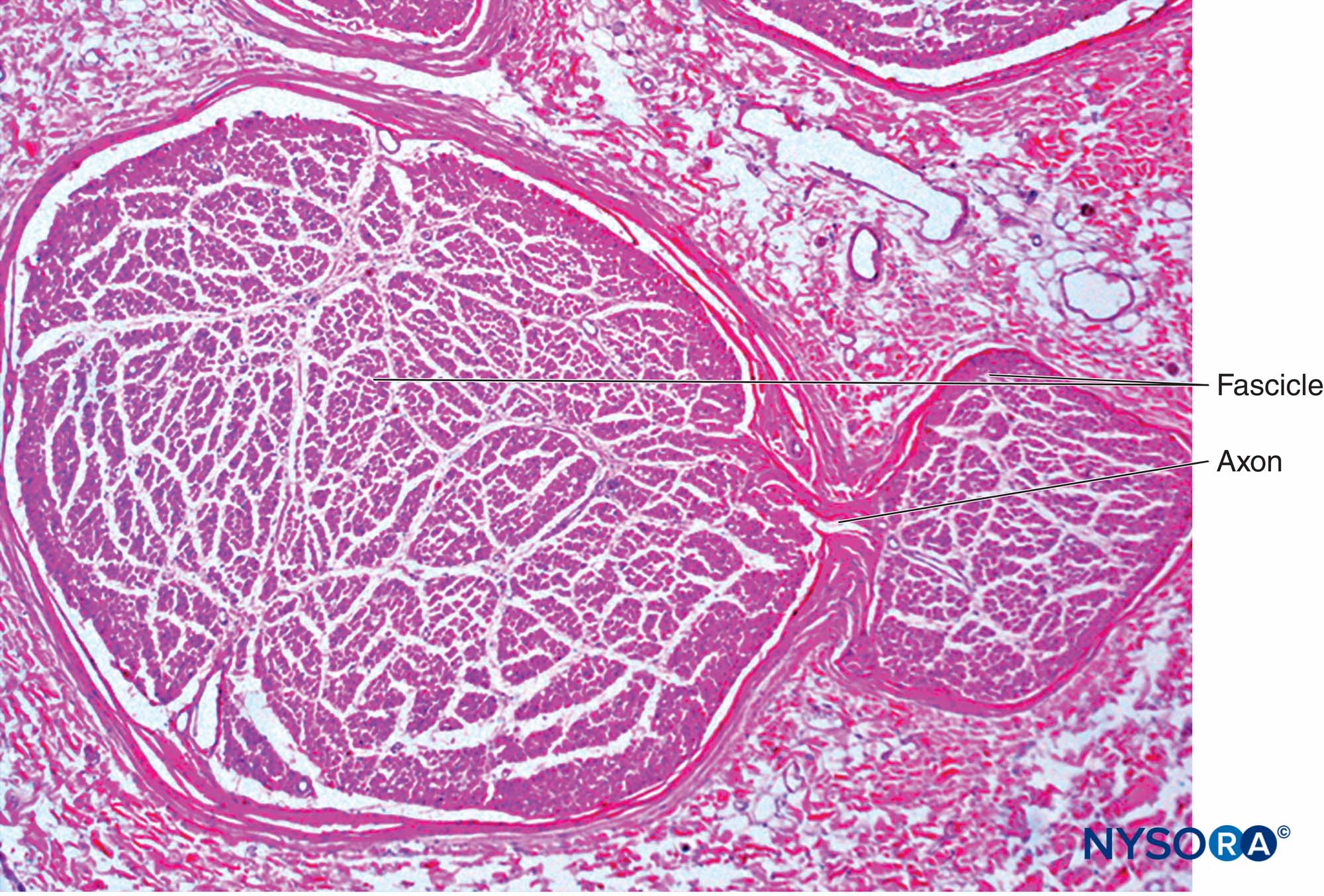
図6 末梢神経内の神経内神経叢。 腕神経叢から得られた、2013つの束の間の軸索からの束間接続。 ヘマトキシリン-エオシン。 (Reina MA、Arriazu R、Collier CBらの許可を得て複製:神経ブロックの実践に臨床的に関連するヒト末梢神経の電子顕微鏡検査。元の実験および実験データに基づく構造的および超構造的レビュー。RevEspAnestesiol Reanim 。60年10月;552(562):XNUMX-XNUMX。)
NYSORAのヒント
各神経の内部で、軸索は、軸索が異なる束を占めることができるように、神経内神経叢を形成します。
坐骨神経の断面では、束は断面積の25%〜75%を占めます(を参照)。 図1 & 3)。 この比率は、神経によって、また同じ神経のレベルによって異なります。 断面積の最大50%は、神経内液や結合間質などの非神経組織で構成されています。 筋束の数は、神経分岐のレベルで増加します。 関節の近くでは、筋肉束はより薄く、より多く、そしてより厚い神経周膜を持っており、それは圧力と伸展に対するより良い保護を与えるかもしれません。
末梢神経の結合組織シート
神経内の結合組織は、神経の血管とリンパ管をサポートおよび保護するように機能します(を参照) 図1 & 2)。 末梢神経の結合組織は、その場所に応じて異なる名前を取ります。 各末梢神経の外側には、コラーゲン組織があります:神経上膜。 神経内のすべての束を取り囲むのは神経周膜です。 束内の個々の神経線維は神経内膜に埋め込まれており、神経周膜によって囲まれた空間を満たします。 末梢神経が分裂し、筋肉束の数が減少するにつれて、結合組織鞘は次第に薄くなります。 たとえば、単束神経では、神経上膜が存在しないか、不規則に分布しているか、神経周膜と統合されているように見えます。 神経を周囲の構造に接続する結合組織は薄く分散しており、一般的な結合組織の特徴と区別できる特徴を失うことがよくあります。
神経内膜
神経内膜(図7 & 8)シュワン細胞を密接に取り囲み、神経周膜によって外部で囲まれた空間を満たします。 神経内膜には、コラーゲン線維、線維芽細胞、毛細血管、およびいくつかの肥満細胞とマクロファージが含まれています。 コラーゲン線維は透過性であり、神経周膜の下および神経線維と血管の周りのゾーンに集中しています。 コラーゲン線維は、有髄神経線維と無髄神経線維の両方を取り囲んでいます。 ただし、小さな有髄線維の周りといくつかの無髄軸索の周りの神経内膜鞘はあまりよく組織化されていません(図9).
図7 複数の小管の形をした神経内膜は、脛骨神経の束を囲んでいます。 電子顕微鏡で観る。 倍率×900。 (レイナの許可を得て複製
MA:局所麻酔および疼痛医学のための機能解剖学のアトラス。 ニューヨーク:スプリンガー; 2015.)
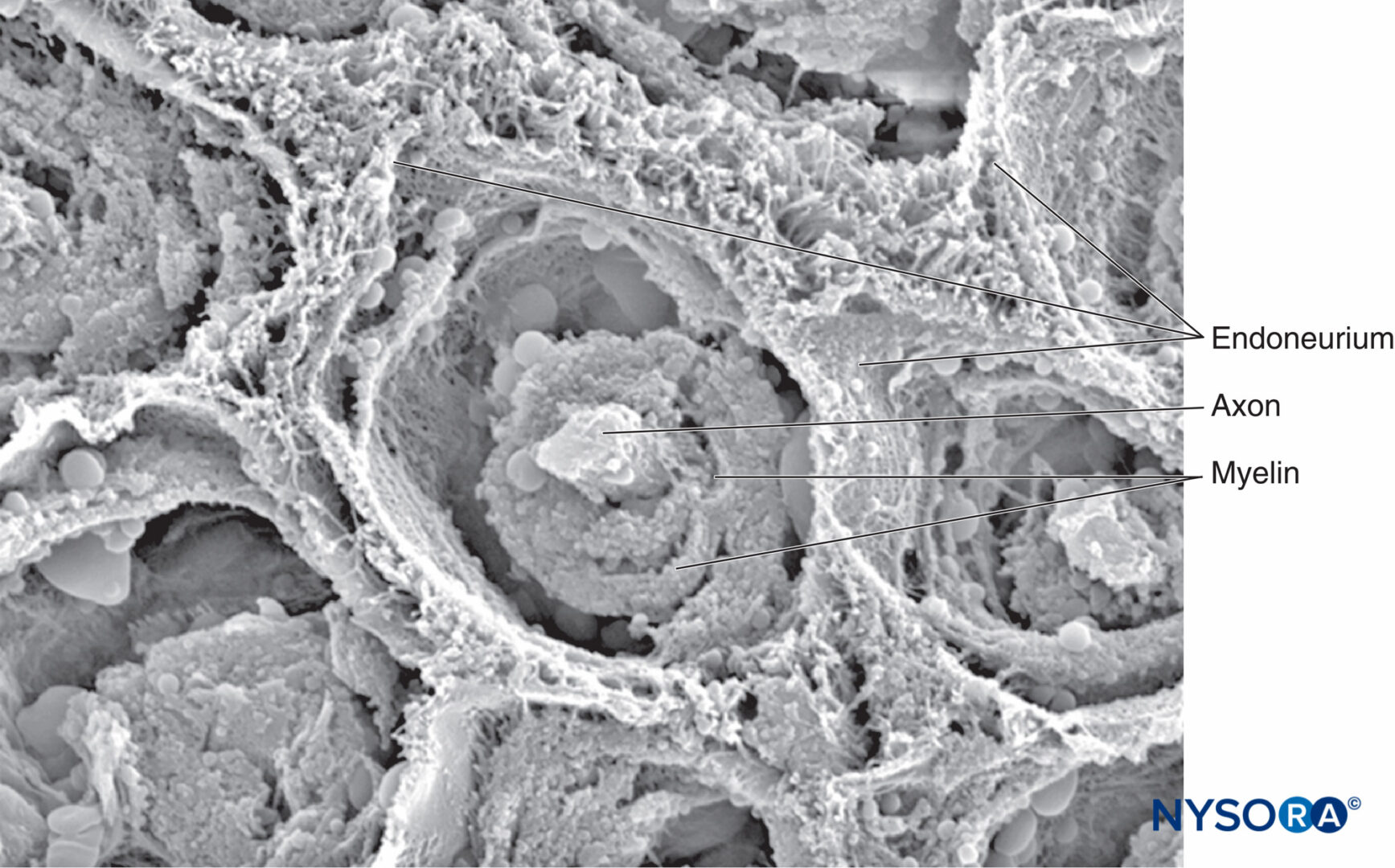
図8 神経内膜は、末梢神経の筋束に有髄軸索を包み込みます。 電子顕微鏡で観る。 倍率×3300。 (Reina MA、Arriazu R、Collier CBらの許可を得て複製:神経ブロックの実践に臨床的に関連するヒト末梢神経の電子顕微鏡検査。元の実験および実験データに基づく構造的および超構造的レビュー。RevEspAnestesiol Reanim 。2013年60月;10(552):562-XNUMX。)
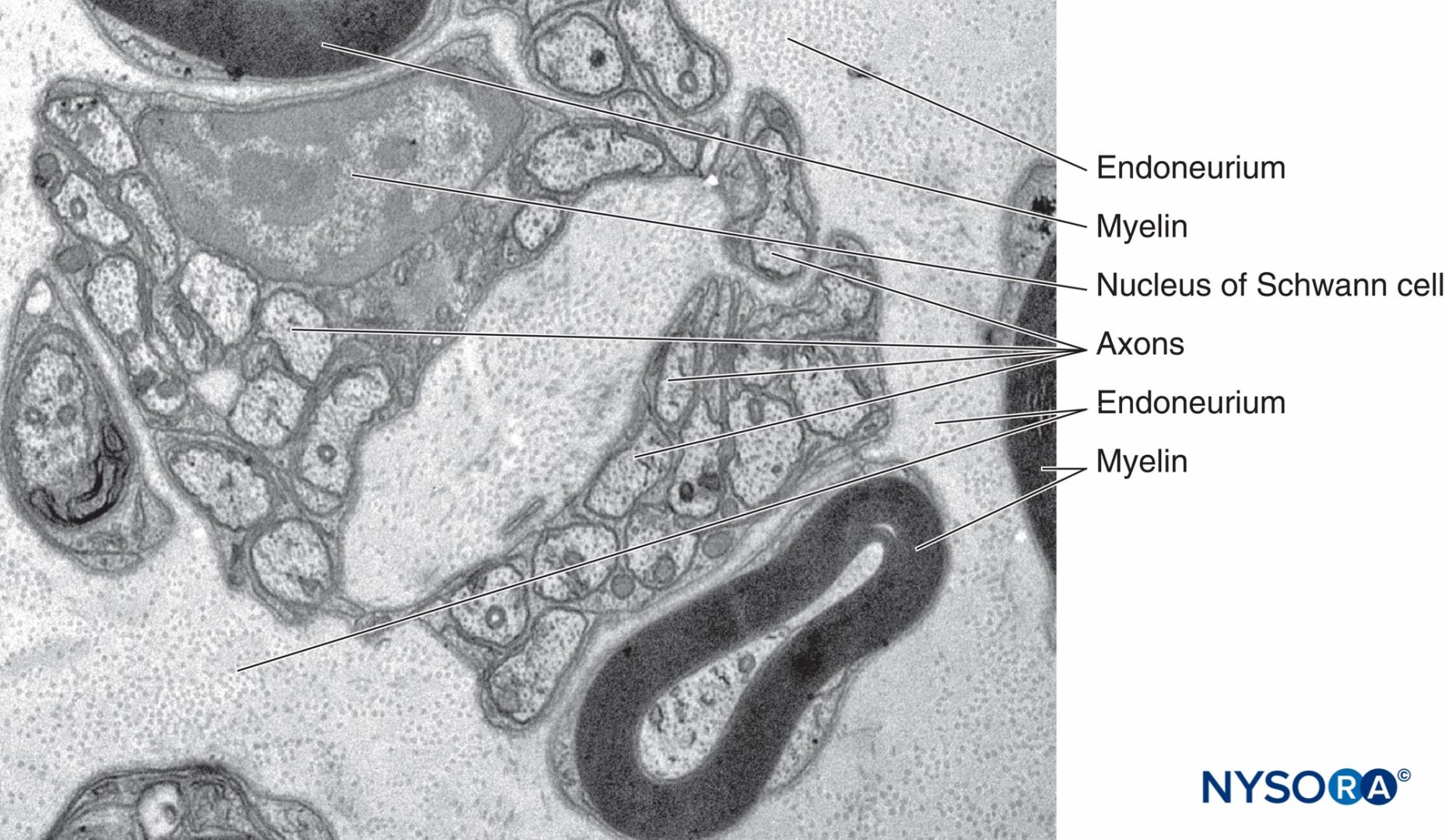
図9 神経内膜に囲まれた無髄および有髄軸索。 透過電子顕微鏡法。 倍率×20000。 (Reina MA、Arriazu R、Collier CBらの許可を得て複製:神経ブロックの実践に臨床的に関連するヒト末梢神経の電子顕微鏡検査。元の実験および実験データに基づく構造的および超構造的レビュー。RevEspAnestesiol Reanim 。2013年60月;10(552):562-XNUMX。
線維芽細胞は、神経内膜の最も豊富な細胞型のXNUMXつです。 彼らは繊維の形成と粉砕された物質の生産に責任があります。 横方向に切断すると、神経内膜線維芽細胞は三角形または長方形のペリカリアを持ちます。 線維芽細胞の外観は、それらの機能的活性によって異なります。 細胞が代謝的に活性である場合、損傷後の成長および組織再生の場合のように、核はより大きく、核小体はより顕著である。 細胞質はまた、より深く染色され、比較的不活性な細胞のわずかに好酸性の細胞質とは対照的に、好塩基性である。 神経上膜のものと同様に、神経内膜の線維芽細胞は基底膜を欠いています。
マスト細胞は、血管の経路に沿って特に多数あります。 肥満細胞の顆粒は水溶性であるため、ヘマトキシリンおよびエオシン染色で日常的に調製された切片では容易に明らかになりません。 適切に固定した後、顆粒はほとんどの塩基性染料で染色され、トルイジンブルーなどの特定の染料の後に異染性になります。 電子顕微鏡写真は、分泌顆粒が膜に結合しており、顆粒マトリックスがさまざまな密度と特徴的ならせん型パターンを持っていることを示しています(図10)。 マクロファージは血管周囲の神経内膜の周りにも頻繁に見られます(図11)。 神経内膜は、シュワン細胞と軸索が位置する内部培地の安定性に貢献しています。 皮膚神経の神経内膜には、深部神経よりも多くのコラーゲン繊維が含まれています。 これはおそらくその保護的役割に関連しています。 神経内コラーゲンは、線維芽細胞よりも9:1多いシュワン細胞に由来すると考えられています。 シュワン細胞は束内細胞の90%を占めますが、線維芽細胞は残りの数の5%未満を占めます。 神経内膜は、神経上膜および神経周膜とともに、緊張下での伸長に対する神経保護に寄与します。 軸索の曲がりくねった軌道は、神経に追加の保護を与えます。 軸索の周りの神経内膜鞘は、 数字7, 8 & 9。 個別に形成された神経内膜層の代わりに、神経内膜はむしろ連続体として現れ、軸索が埋め込まれているいくつかの小管を形成します。
")
図10 脛骨神経からの束内の肥満細胞。 透過電子顕微鏡法。 倍率×7000。 (Reina MAの許可を得て複製:局所麻酔および疼痛医学のための機能解剖学のアトラス。ニューヨーク:Springer; 2015年。)

図11 透過型電子顕微鏡で見られる束内のマクロファージ。 倍率×7000。
NYSORAのヒント
神経内膜はシュワン細胞を取り囲み、神経周膜内の空間を満たします。
神経周膜
各筋束は、結合組織鞘である神経周膜に囲まれています。 神経周膜は、コラーゲンの層によって分離された平らな細胞の同心層で構成されています(図12 〜へ 16)。 神経周膜細胞層の数は、筋肉束のサイズによって異なります。 大きな神経束の周りには8〜16の同心層が存在する可能性がありますが、神経周膜細胞の単層が小さな遠位束を囲んでいます。 より大きな末梢神経では、同心細胞層は、神経上膜のものと同様に、縦方向に配置されたコラーゲン線維の層と交互になっています。 コラーゲン線維は神経上膜よりも細く、その間に散在する弾性線維はごくわずかです。 神経周膜細胞は、両側にかなり密集している可能性のある基底膜を持っています。 ヘミデスモソームとして知られる部位では、神経周膜細胞の原形質膜が基底膜に強く付着します。
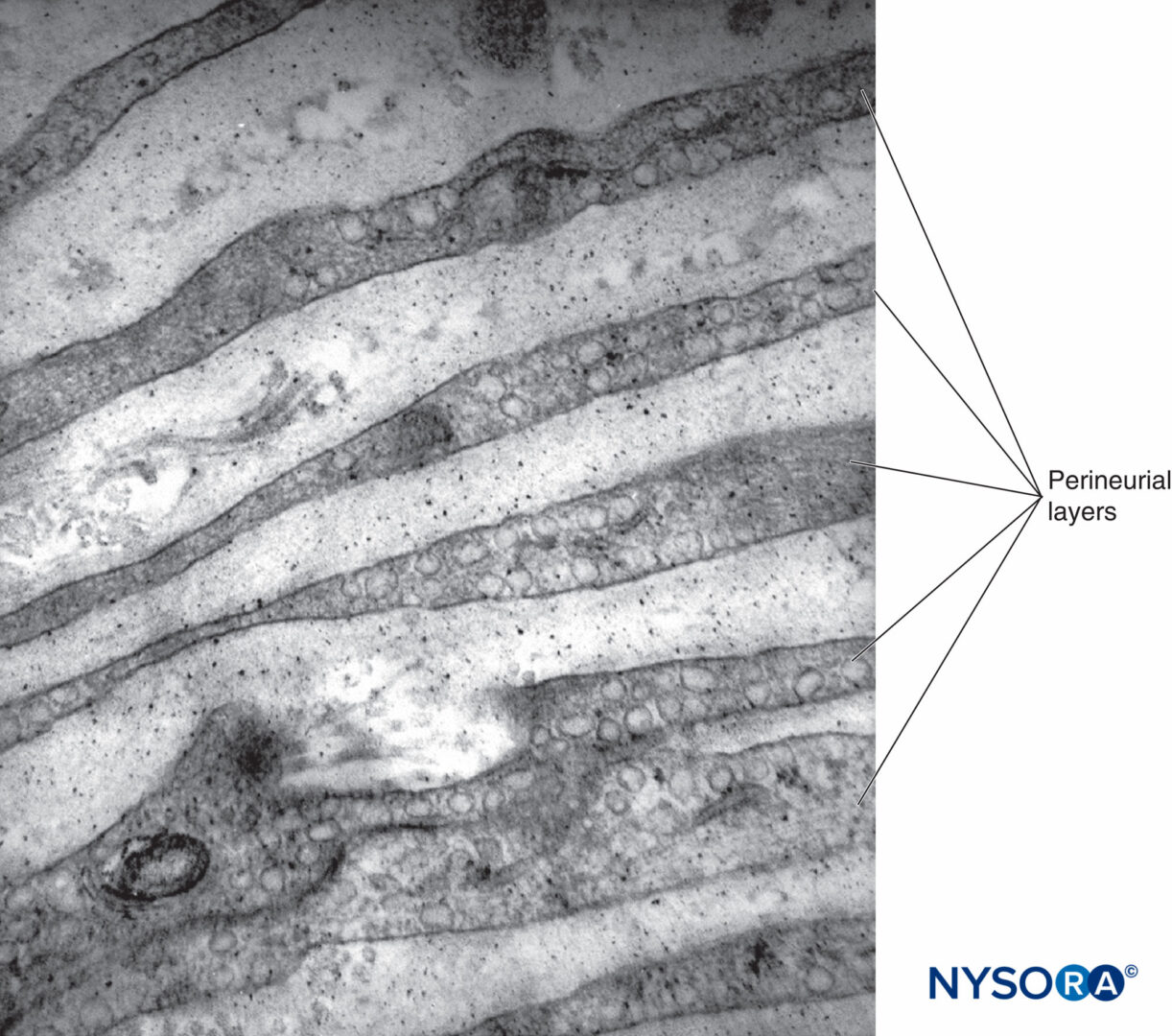
図12 同心神経周膜層。 透過電子顕微鏡法。 倍率×30,000。 (Reina MA、Arriazu R、Collier CBらの許可を得て複製:神経ブロックの実践に臨床的に関連するヒト末梢神経の電子顕微鏡検査。元の実験および実験データに基づく構造的および超構造的レビュー。RevEspAnestesiol Reanim 。2013年60月;10(552):562-XNUMX。)
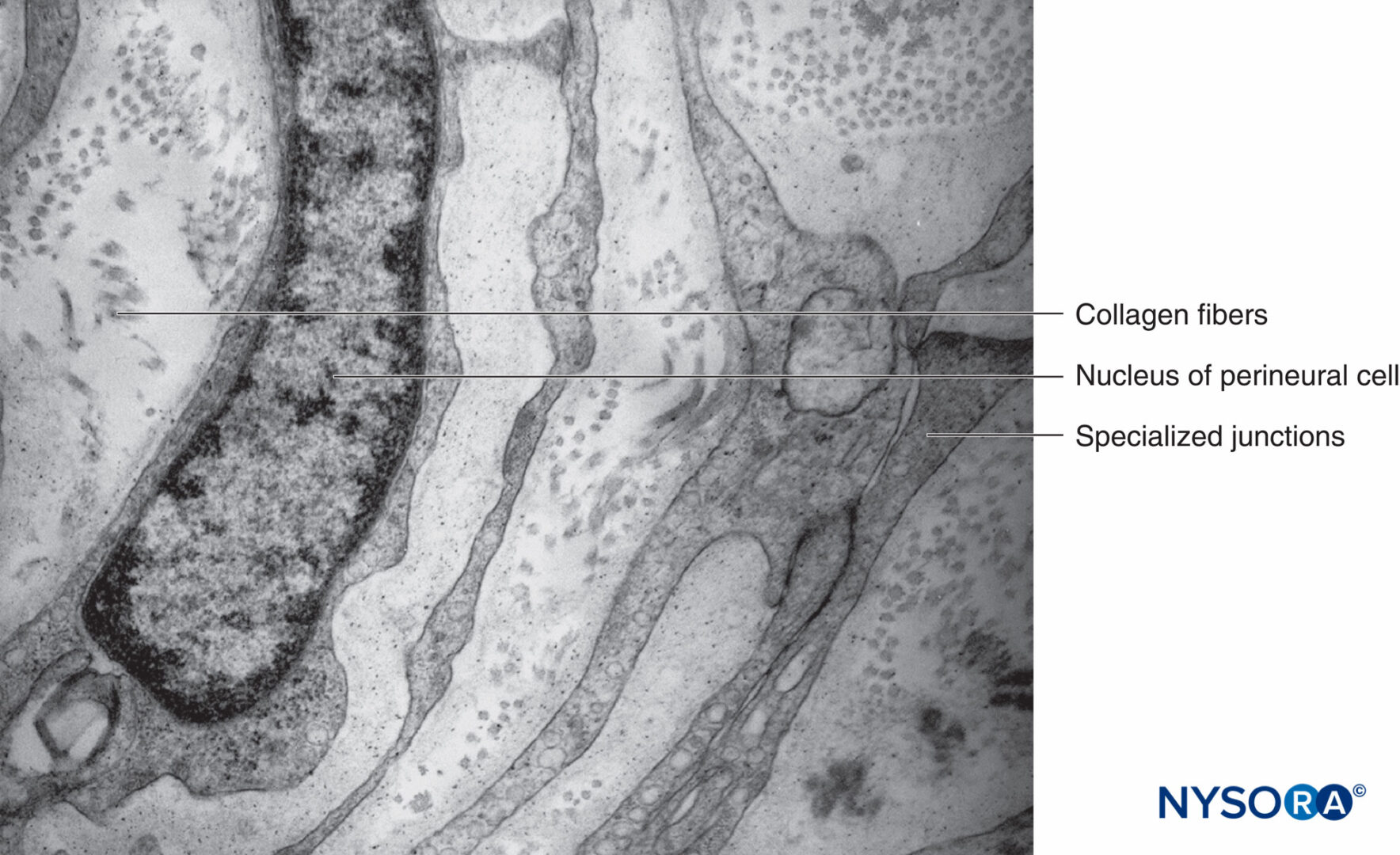
図13 神経周膜層と特殊な接合部。 透過電子顕微鏡法。 倍率×20,000。 (Reina MA、Arriazu R、Collier CBらの許可を得て複製:神経ブロックの実践に臨床的に関連するヒト末梢神経の電子顕微鏡検査。元の実験および実験データに基づく構造的および超構造的レビュー。RevEspAnestesiol Reanim 。2013年60月;10(552):562-XNUMX。)
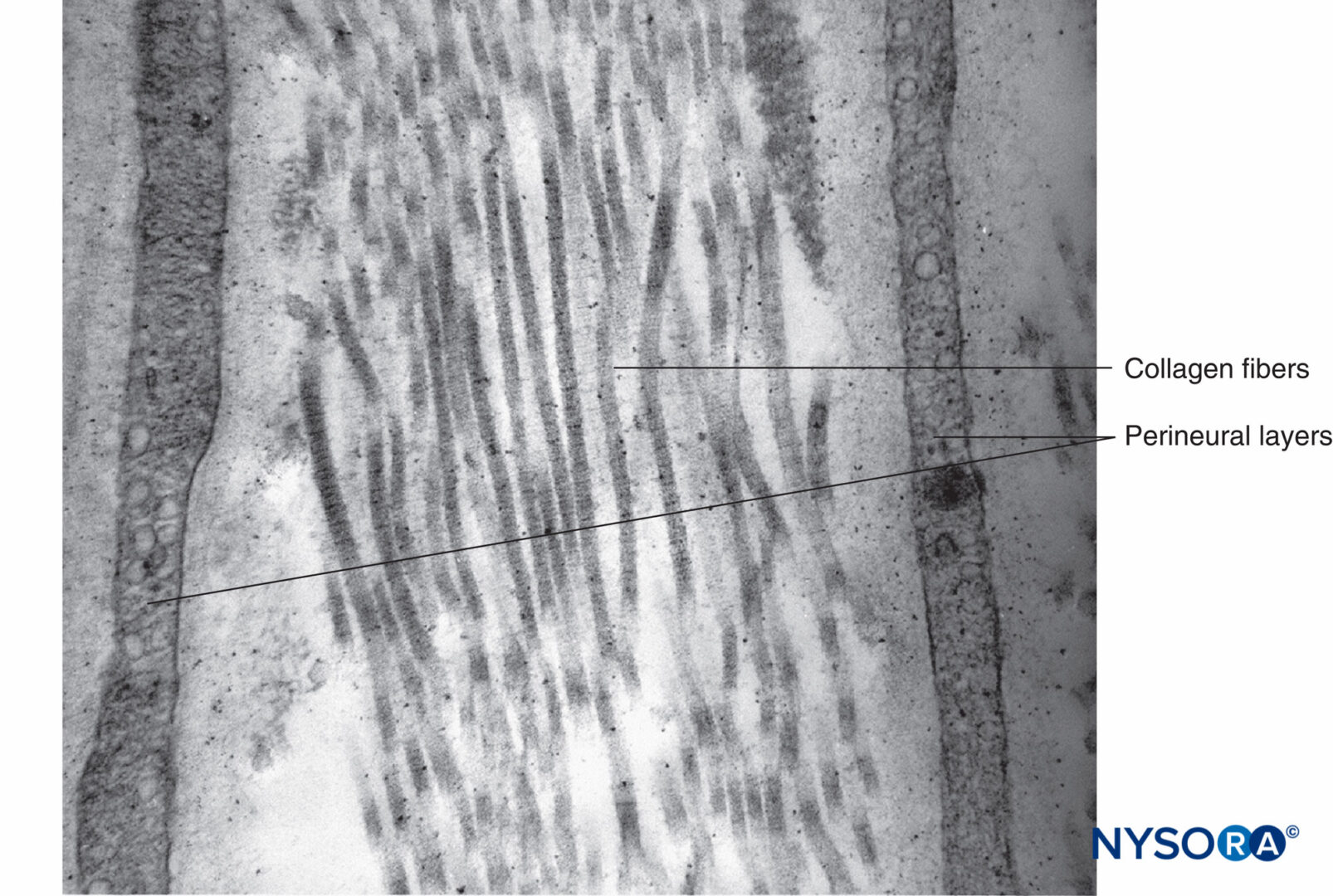
図14 神経周膜層の間のコラーゲン線維。 透過電子顕微鏡法。 倍率×30,000。 (Reina MAの許可を得て複製:局所麻酔および疼痛医学のための機能解剖学のアトラス。ニューヨーク:Springer; 2015年。)
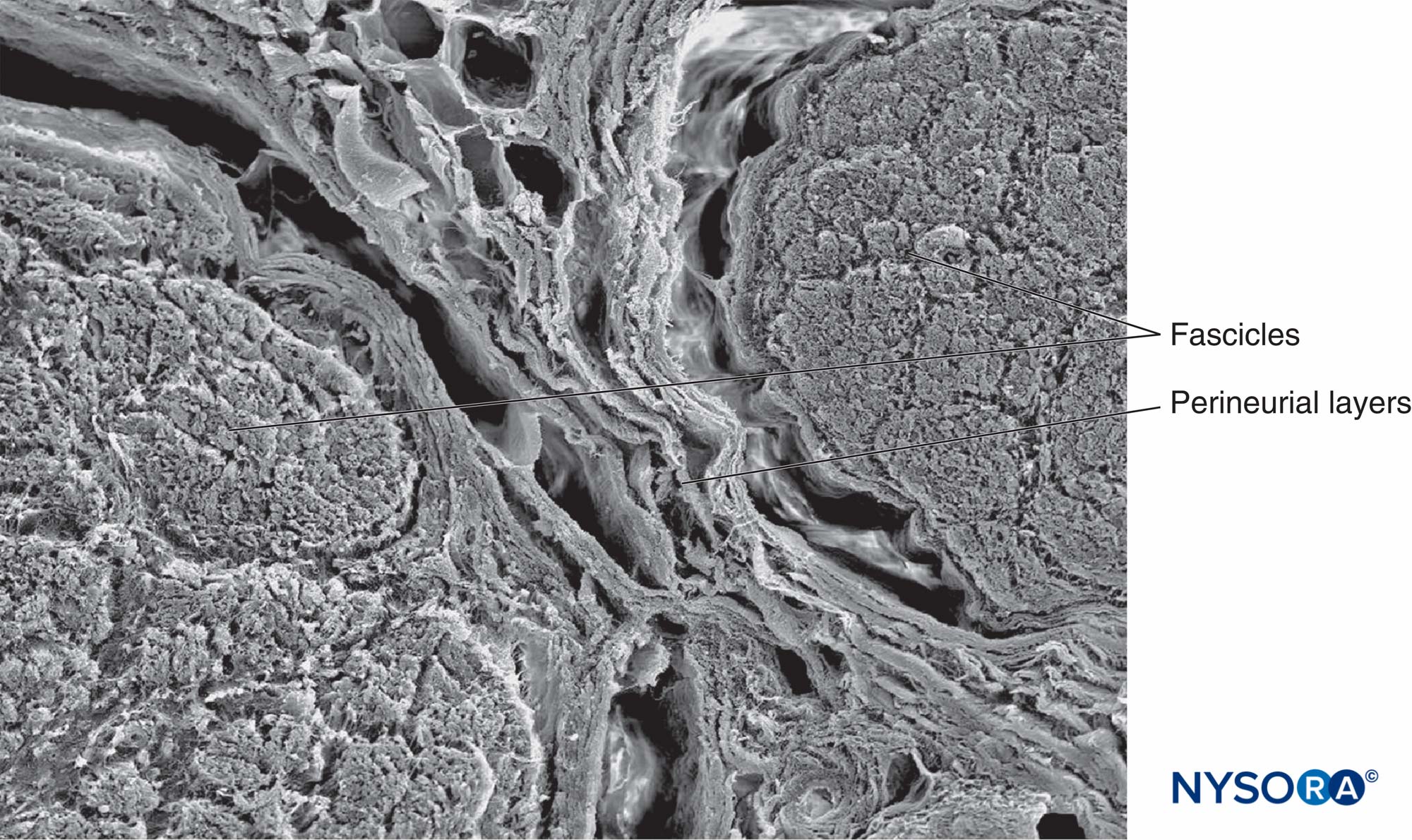
図15 神経周膜と筋肉束:神経周膜層の150次元的特徴。 電子顕微鏡で観る。 倍率×2013。 (Reina MA、Arriazu R、Collier CBらの許可を得て複製:神経ブロックの実践に臨床的に関連するヒト末梢神経の電子顕微鏡検査。元の実験および実験データに基づく構造的および超構造的レビュー。RevEspAnestesiol Reanim 。60年10月;552(562):XNUMX-XNUMX。)
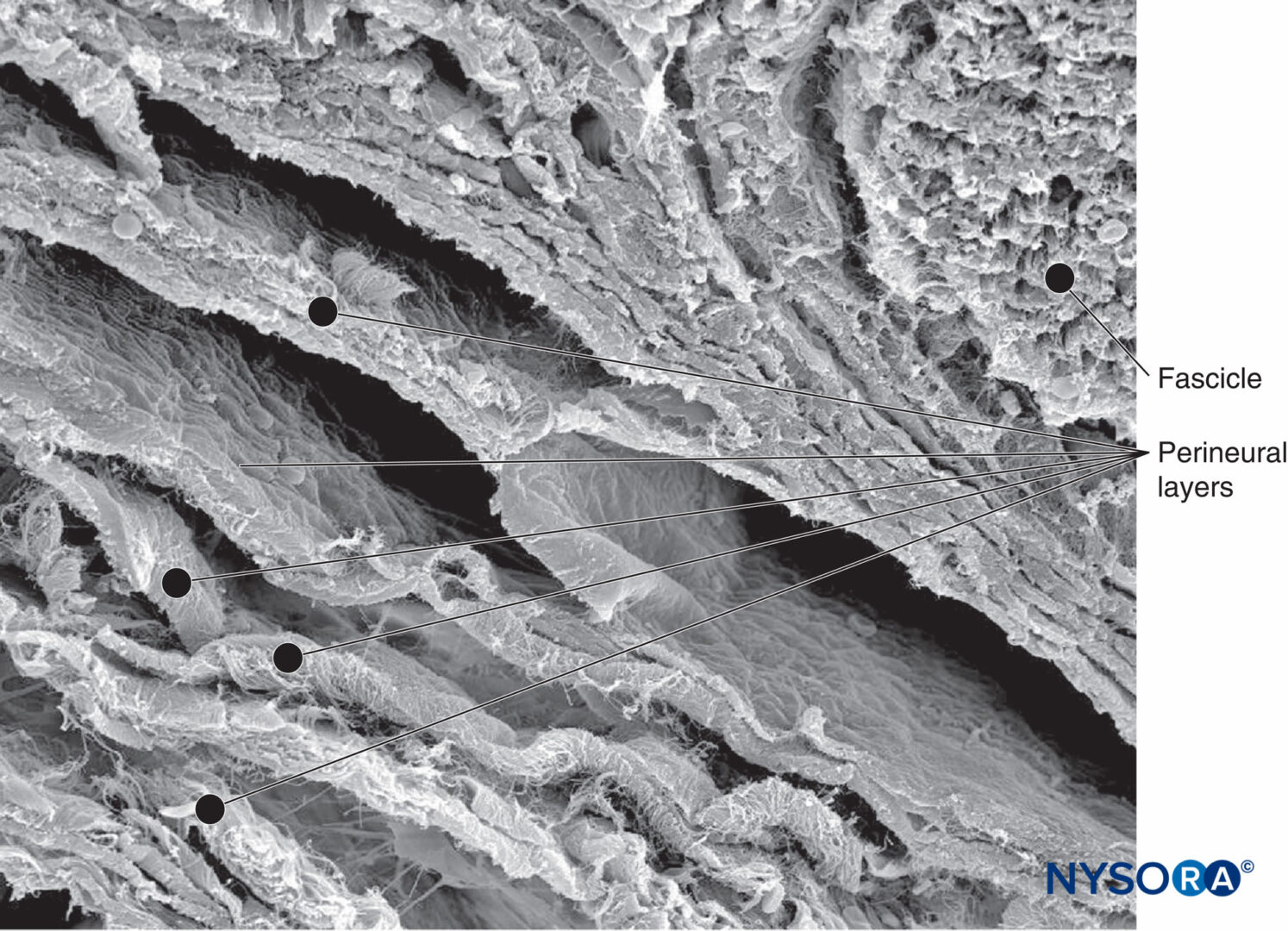
図16 神経周囲層の500次元画像。 電子顕微鏡で観る。 倍率×2013。 (Reina MA、Arriazu R、Collier CBらの許可を得て複製:神経ブロックの実践に臨床的に関連するヒト末梢神経の電子顕微鏡検査。元の実験および実験データに基づく構造的および超構造的レビュー。RevEspAnestesiol Reanim 。60年10月;552(562):XNUMX-XNUMX。)
電子顕微鏡では、神経周囲細胞は、少量の小胞体、フィラメント、および多数のエンドサイトーシス小胞を含む細胞質の薄いシートとして見られます。 神経周膜の同じ層内の隣接する細胞間の密着結合およびギャップ結合も観察されます。 同様の密着結合は、細胞が近接している場合、神経周膜の連続する層の間に現れることもあります。 神経周膜の内層の密着結合と神経内膜毛細血管の密着結合は、血液神経関門構造を形成します(図17 & 18)。 血液脳関門星状細胞は、血液と脳の間の化合物の流れを調節するのに役立つため、血液脳関門は血液脳関門と同等ではありません。 神経周囲細胞は代謝的に活性であり、それらの細胞質にはATPase(アデノシントリホスファターゼ)、5-ヌクレオチダーゼなどの酵素が含まれています。 これらの細胞は、おそらく神経細胞の周りの電解質とブドウ糖のバランスを維持する役割を果たしています。
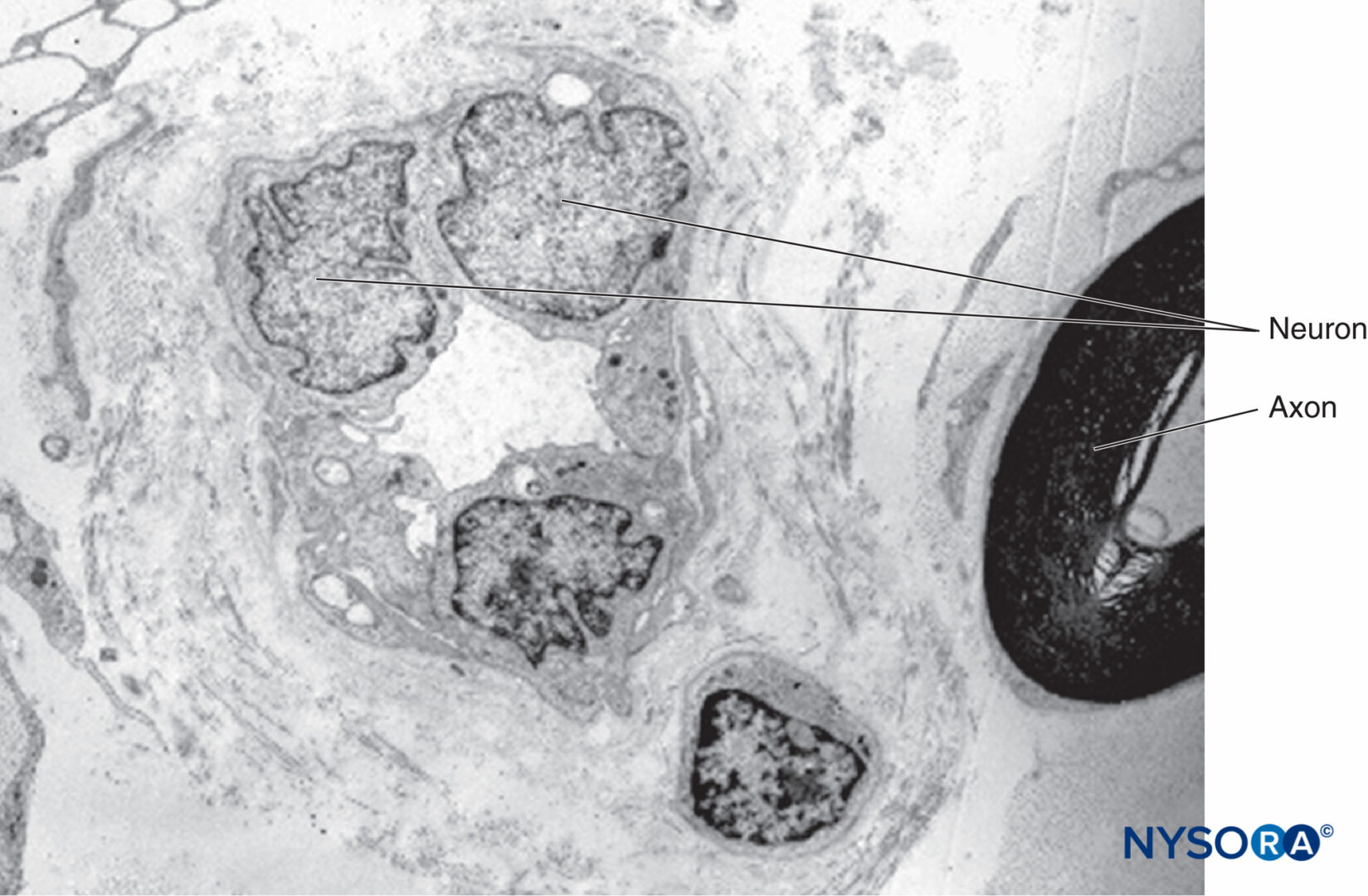
図17 末梢神経束内の神経内膜と毛細血管。 透過電子顕微鏡法。 倍率×3000。 (Reina MA、Arriazu R、Collier CBらの許可を得て複製:神経ブロックの実践に臨床的に関連するヒト末梢神経の電子顕微鏡検査。元の実験および実験データに基づく構造的および超構造的レビュー。RevEspAnestesiol Reanim 。2013年60月;10(552):562-XNUMX。)
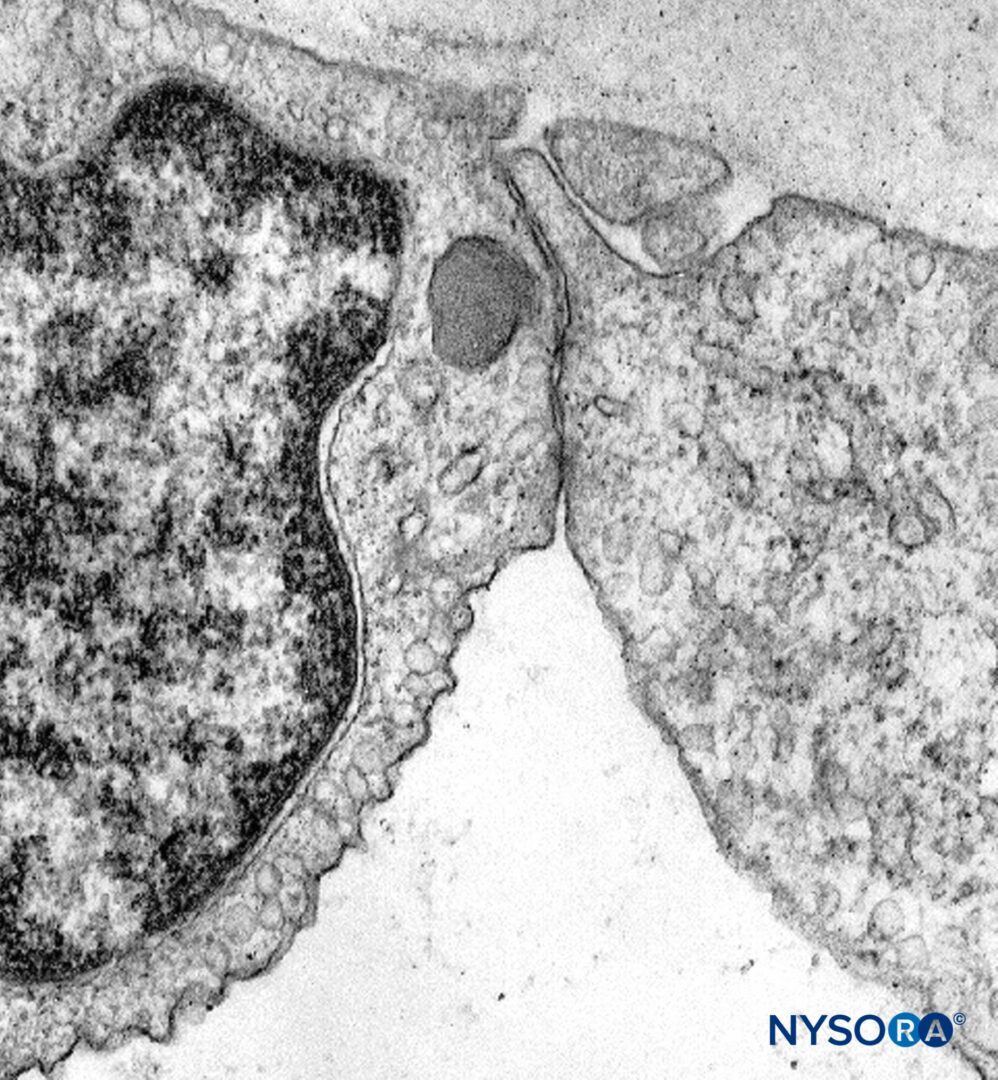
図18 束内毛細血管からの内皮細胞。 透過電子顕微鏡法。 倍率×20,000。 (Reina MA、LópezA、Villanueva MCらの許可を得て複製:末梢神経の血液神経関門。RevEspAnestesiolReanim。2003年50月;2(80):86-XNUMX。)
神経周膜は管状のラッピングを形成し、これにより筋束内で軸索の動きが可能になります。 神経上膜の厚さは1から100μmの間で変化します。 神経内の筋束の数が増えると、神経周膜の厚さは一般に減少します。 たとえば、正中神経の軌道に沿って、神経上膜は腋窩よりも手首で比例して厚く見えます。 神経周膜が存在せず、神経上膜が神経内膜と接触する領域はXNUMXつあります。神経終末、血管の周囲、および細網線維が神経周膜を貫通する領域です。
神経周膜の役割は、束内圧を維持し、バリア効果に寄与することです。 神経周膜にかかる圧力は神経内膜に伝達され、最終的には神経線維(軸索)に伝達されます。 神経周膜は、神経分岐点の周りで厚さが増し、追加の保護を提供します。 神経周膜はまた、感染および炎症反応の拡大を制限するのに保護的である可能性があります。 たとえば、神経周膜が無傷の神経が感染領域を横切ると、神経は一般に神経周膜の肥厚によって反応します。 逆に、神経周膜が無傷でない場合、感染は神経束全体に容易に広がります。 ただし、神経上膜の損傷は、軸索の安全性を同程度に損なうものではありません。 Söderfeltは、神経上膜のバリア効果が、虚血状態で死後22時間まで維持されることを実証しました。 オルソンは、神経損傷後の生体内での神経バリア効果の喪失を研究しました。 彼はまた、損傷後2日から30日の間に効果の回復を示しました。
NYSORAのヒント
- 神経内の束は神経周膜に囲まれており、神経周膜は貫通や過度の伸展による損傷に対する構造的保護をもたらします。
- 血液脳関門は、神経周膜の内層の密着結合と神経内膜毛細血管の密着結合によって形成されます。
神経上膜
神経上膜の最も外側の鞘は、神経束を結合する適度に密な結合組織で構成されています(図3, 19, 20)。 神経上膜は、特に皮下組織において、末梢神経を取り巻く脂肪組織と融合します。 上神経組織の量は神経に沿って変化し、関節の周りにより豊富になります。 神経上膜の厚さは、神経によって、また同じ神経の場所によって異なります。 たとえば、神経上膜の平均的な厚さは、肘のレベルでは尺骨神経の22%、臀部のレベルでは坐骨神経の88%です。
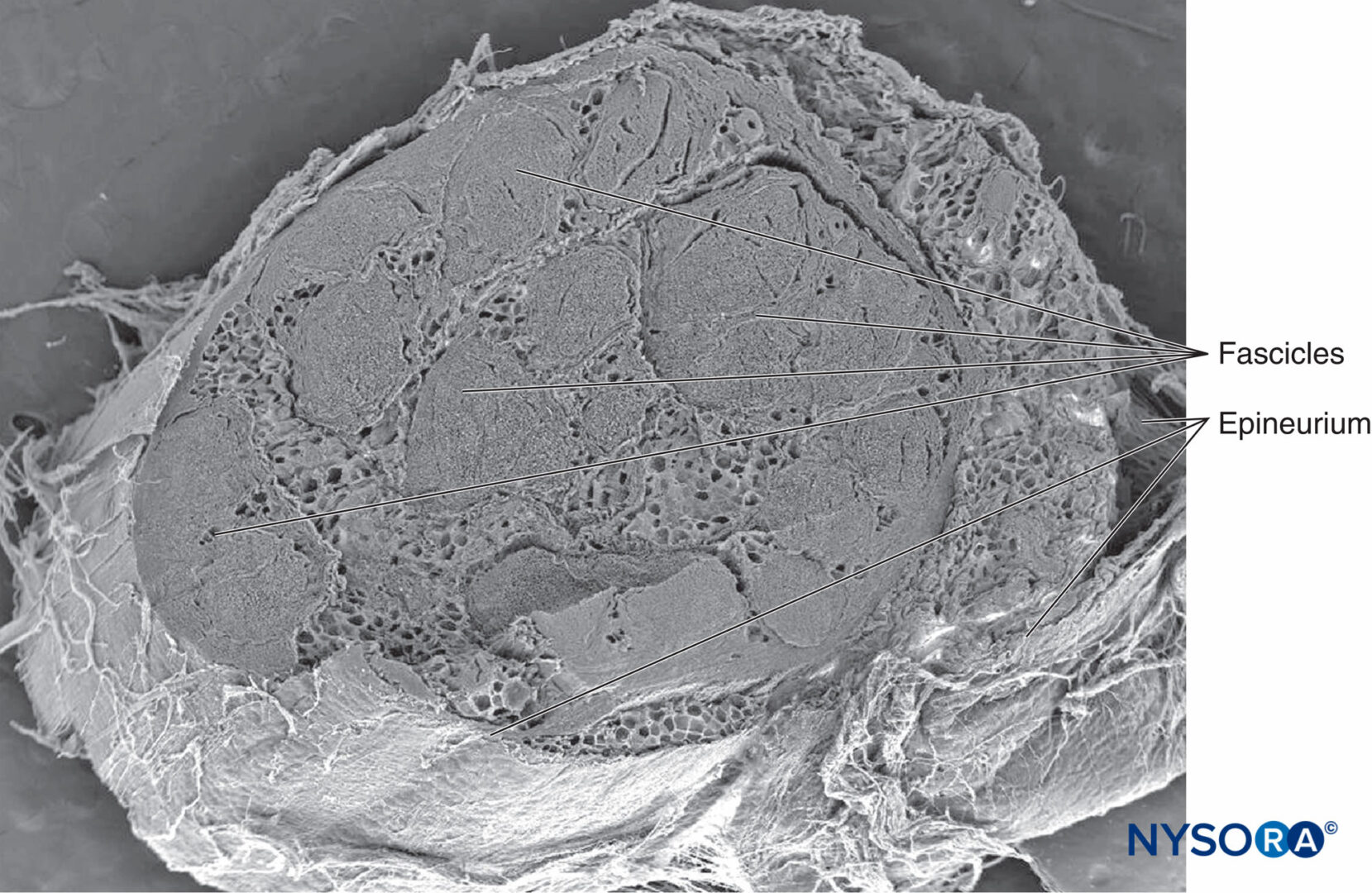
図19 ヒト脛骨神経の神経上膜。 電子顕微鏡で観る。 倍率×20。 (Reina MA、Arriazu R、Collier CBらの許可を得て複製:神経ブロックの実践に臨床的に関連するヒト末梢神経の電子顕微鏡検査。元の実験および実験データに基づく構造的および超構造的レビュー。RevEspAnestesiol Reanim 。2013年60月;10(552):562-XNUMX。)
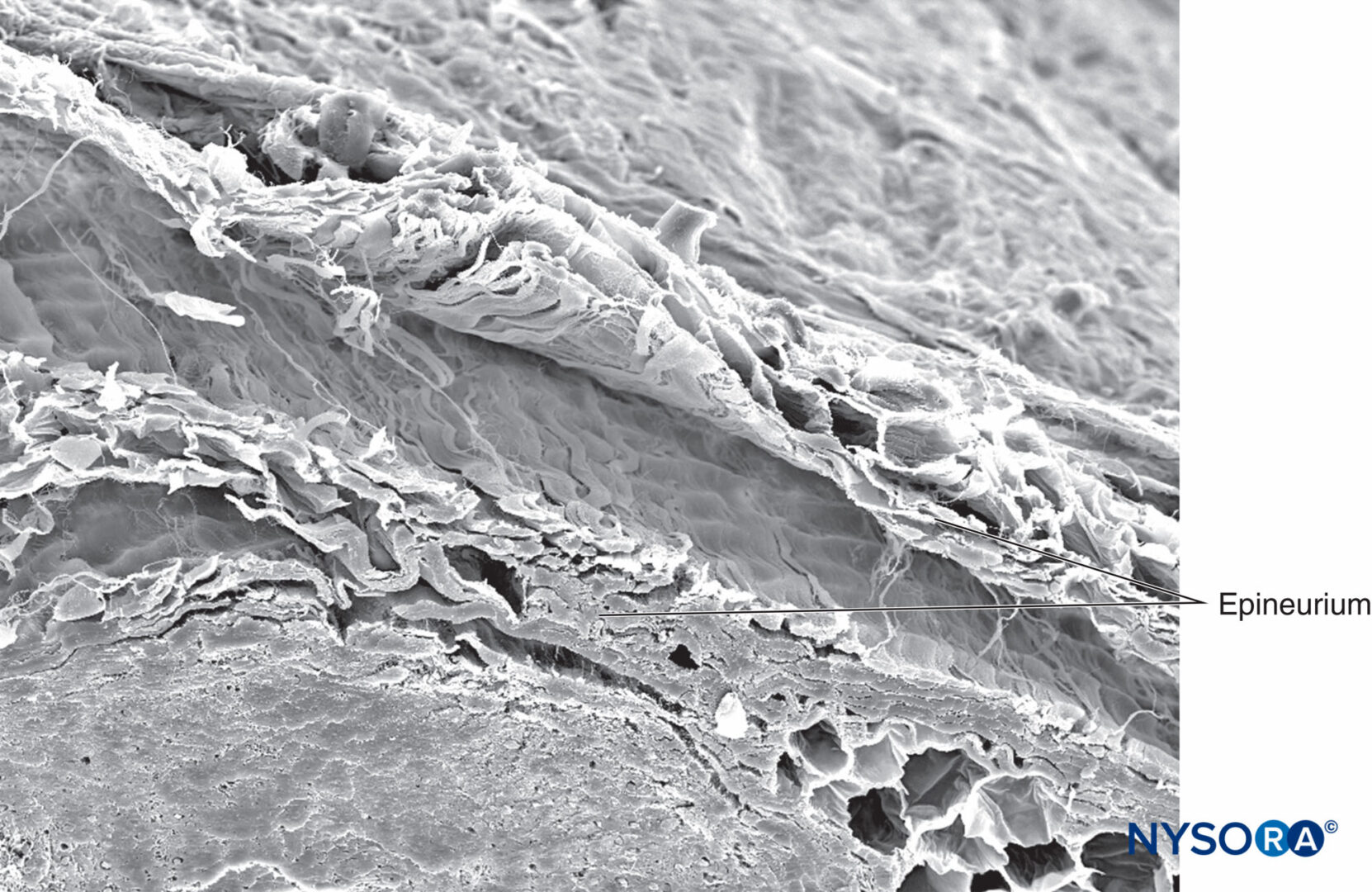
図20 ヒト脛骨神経の神経上膜。 電子顕微鏡で観る。 倍率×180。 (Reina MA、Arriazu R、Collier CBらの許可を得て複製:神経ブロックの実践に臨床的に関連するヒト末梢神経の電子顕微鏡検査。元の実験および実験データに基づく構造的および超構造的レビュー。RevEspAnestesiol Reanim 。2013年60月;10(552):562-XNUMX。)
一般に、神経上膜は神経の断面積の30%から75%を占めます。 神経上膜の割合は、神経束の数が増えるにつれて、より大きな神経でより高くなります。 ただし、神経上膜は単束神経の周囲と神経終末には存在しません。神経上膜には、脂肪細胞、線維芽細胞、結合組織線維、マスト細胞、小血管とリンパ管、および血管を神経支配する小神経線維が含まれています。 神経上膜は透過性の構造であり、その線維芽細胞は体の他の場所の線維芽細胞と超微細構造的に同一です。 神経上膜全体に散在する線維芽細胞は、この層の最も顕著な成分である神経上膜コラーゲンを形成します。 コラーゲンはほとんどの酸性染料で染色されるタンパク質であるため、ヘマトキシリン-エオシンで染色された製剤では、コラーゲン繊維がエオシンで弱いピンク色に変わります。 電子顕微鏡下では、成熟コラーゲンの繊維は頻繁にクロスバンディングを起こします。 弾性繊維も存在し、これらはコラーゲン繊維よりもかなりコンパクトです。 それらは、ヘマトキシリンとエオシンで染色されたセクションで弱いピンクに染色され、オルセインで茶色に染色され、レゾルシン-フクシンで青紫色に染色されます。 電子顕微鏡写真では、エラスチン繊維は通常、周辺部でより染色され(暗く)見え、細いエラスチンフィラメントを含む物質に埋め込まれています。一部の神経の上膜には、坐骨神経の場合と同様に、かなりの量の脂肪が含まれています。 ただし、総腓骨神経と脛骨神経は坐骨神経よりも脂肪が少なく、通常、前者は後者よりも脂肪が少なくなっています。
顕微鏡で見ると、神経内脂肪細胞はハニカムに似ており、固定プロセス中は脂肪の溶解により液胞が空になります(図21)。 マスト細胞は結合組織全体に分布しており、多くの場合、小さな血管の近くにあります。 末梢神経を供給するVasanervorumは、局所動脈の枝から発生します。 これらの動脈からの枝は神経上膜に入り、血管叢を形成します(図22 & 23)。 神経叢から、血管は神経周膜を貫通し、細動脈および毛細血管として神経内膜に入ります。 いくつかの束からなる神経では、動脈、静脈、リンパ管が縦方向に走り、神経束と平行に走っています。
神経上膜はまた、その軌道に沿って縦方向の「波動」を投射し、特に四肢に神経支配を供給する神経に弾力性を提供します。
:397-402。)")
図21 坐骨神経の束間組織の脂肪細胞。 電子顕微鏡で観る。 倍率×400。 (Reina MA、LópezA、DeAndrésJAの許可を得て複製:末梢神経内の脂肪組織。人間の坐骨神経の研究。RevEspAnestesiolReanim。2002Oct; 49(8):397-402。)
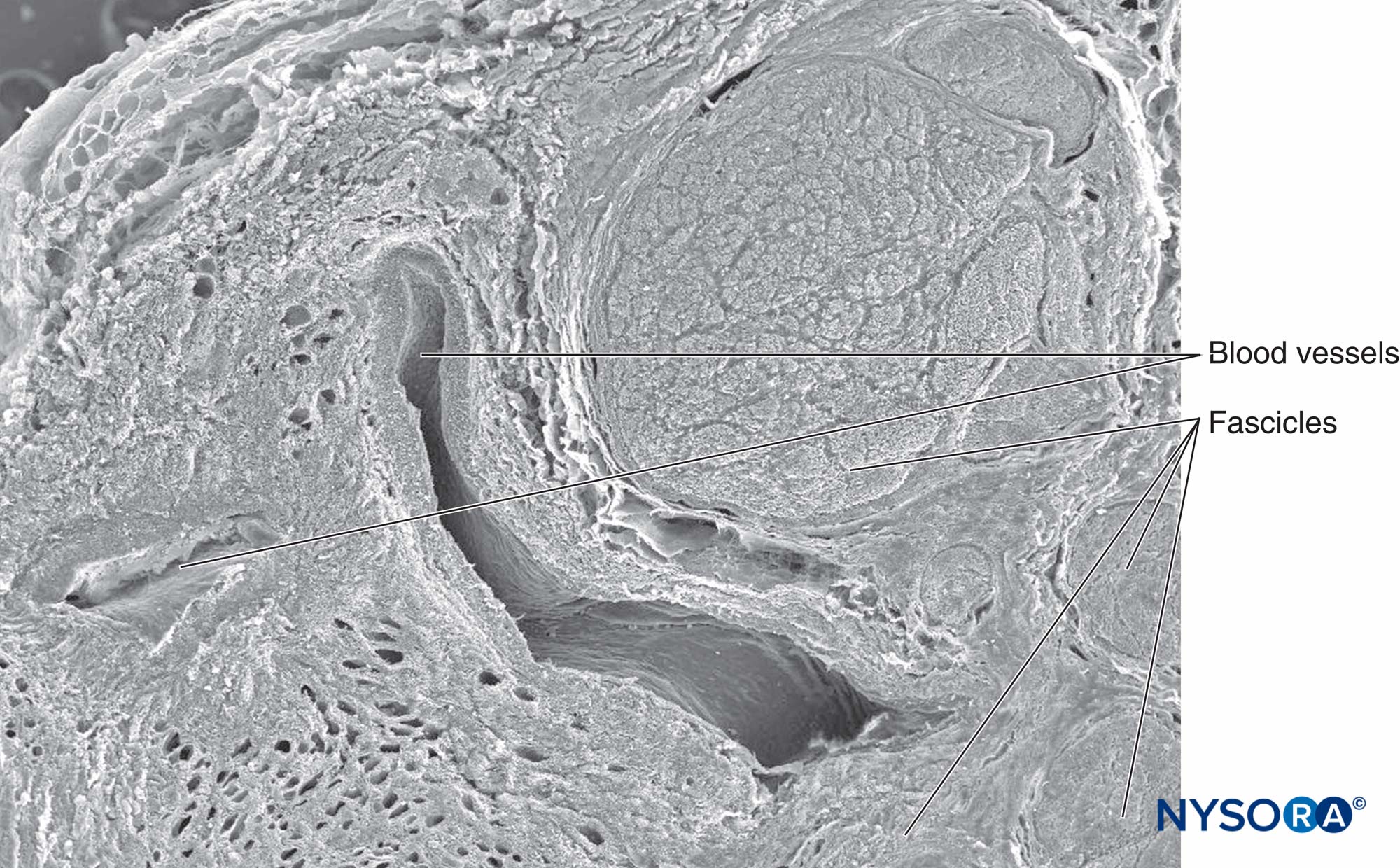
図22 坐骨神経の束間組織および血管。 電子顕微鏡で観る。 倍率×50。 (Reina MA、LópezA、DeAndrésJAの許可を得て複製:末梢神経内の脂肪組織。人間の坐骨神経の研究。RevEspAnestesiolReanim。2002Oct; 49(8):397-402。)
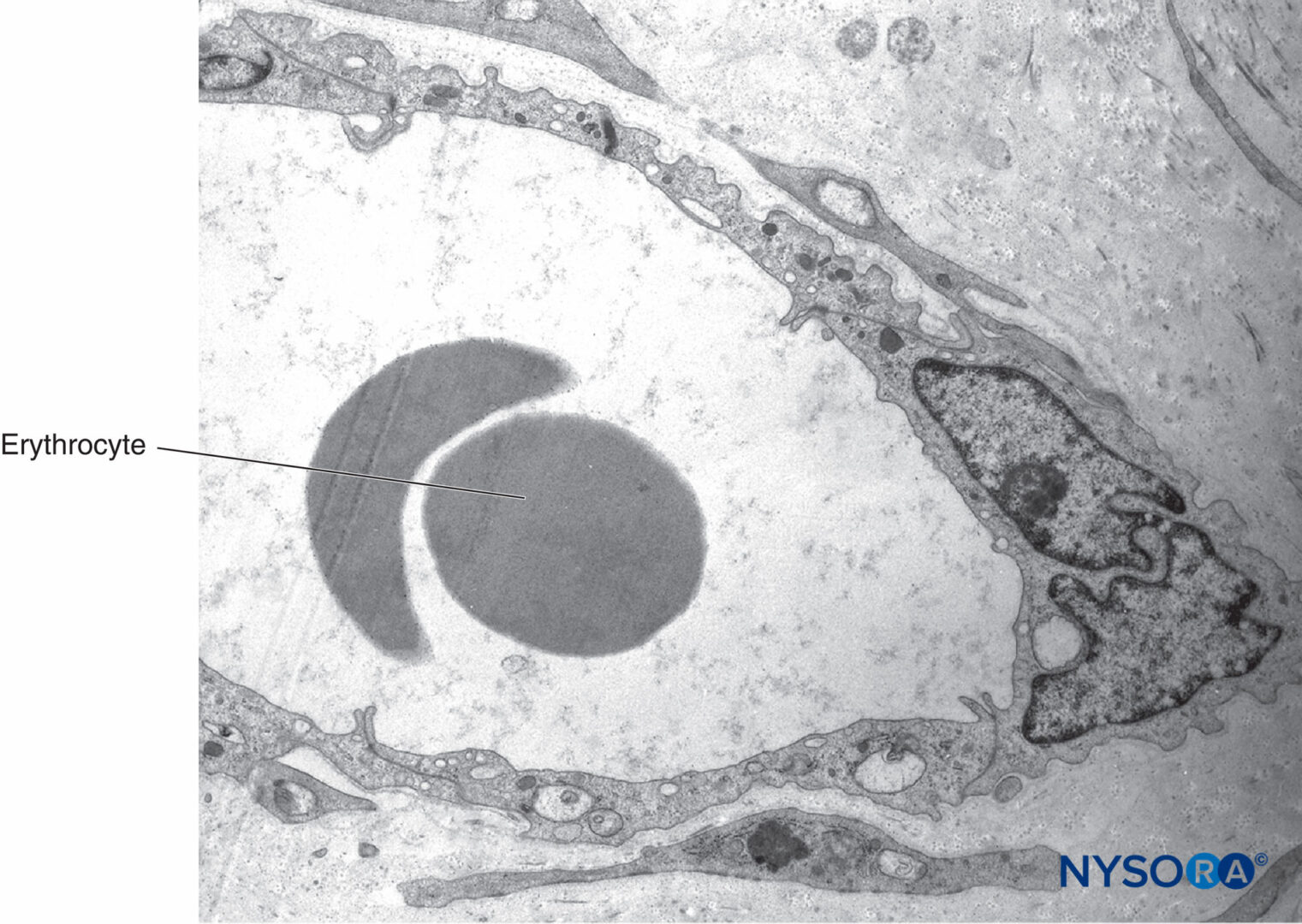
図23 坐骨神経の束間組織内の血管。 透過電子顕微鏡法。 倍率×7000。 (Miguel Angel Reina、MDの許可を得て使用。)
NYSORAのヒント
- 神経上膜は末梢神経の最も外側の鞘です。
- 神経上膜は透過性であり、神経束を結合する適度に密な結合組織で構成されています。
- 神経上膜には、脂肪細胞、線維芽細胞、結合組織線維、肥満細胞、小リンパ管、血管、および血管を神経支配する小神経線維が含まれています。
傍神経鞘および一般的な上腹部鞘
遠位末梢神経の全体的な解剖学的記述は、軸索(神経内膜)、神経束または軸索の束(神経周膜)、および単一の末梢神経(神経上膜)を囲む各結合層を正確に識別しますが、結合組織が複数の神経に結合する場合、これはより複雑になります。 この例は、膝窩の坐骨神経です。 傍神経節、傍神経鞘、一般的なエピネウラル鞘、結膜神経、または外膜などのさまざまな用語は、同じ結合組織を指すために交換可能に使用されます。 筋肉束の単一のグループによって形成される小さな神経は、乏しい量の脂肪組織とともに各筋肉束を囲む神経周膜の層を特徴としています。 コラーゲン線維で構成される神経上膜として知られる結合組織が神経を囲んでいます。 特定の技術により、EMA(上皮膜抗原)陽性の染色法とマッソントリクロームおよびEMA陰性染色によるコラーゲンの染色法を使用して神経周膜の同定が可能になります。
同様に、染色技術は、さまざまな数の束のグループが存在する坐骨神経などのより複雑な神経の構造の識別に役立ちます。 これらのタイプの神経では、EMA染色により、神経束のグループを囲む神経上膜に通常存在するコラーゲン線維で構成される結合組織とは対照的に、神経周膜が各神経束を囲むことが明らかになります(マッソントリクロミック染色で検出)。 ますます近位の位置にある坐骨神経などの複雑な神経構造の顕微鏡分析は、これらの神経構造内の神経枝が、物理的な枝の分割が実現する前でさえ、それぞれの気管層によって分割されているように見えることを示しています。 プレクサスと末端部位の両方の各末梢神経は、脂肪組織の同心クラスターに囲まれています。これは、神経の分裂の直前に現れます(図24 & 25)。 脂肪組織は、その側枝または末端の枝に沿って伸びています。 脂肪組織の量と形状は神経に沿って変化し、徐々に同心円状の輪郭を失い、不均一に分布するようになります。 神経上膜と同様に、神経成分とその間の脂肪組織を一緒に包むコラーゲン層は、異なる著者によって、傍神経膜、傍神経鞘、一般的なエピネウラル鞘、結膜神経または外膜と呼ばれています。
")
図24 膝窩のレベルでの坐骨神経とその周囲の傍神経節。 ヘマトキシリン-エオシン。 (Reina MAの許可を得て複製:局所麻酔および疼痛医学のための機能解剖学のアトラス。ニューヨーク:Springer; 2015年。)
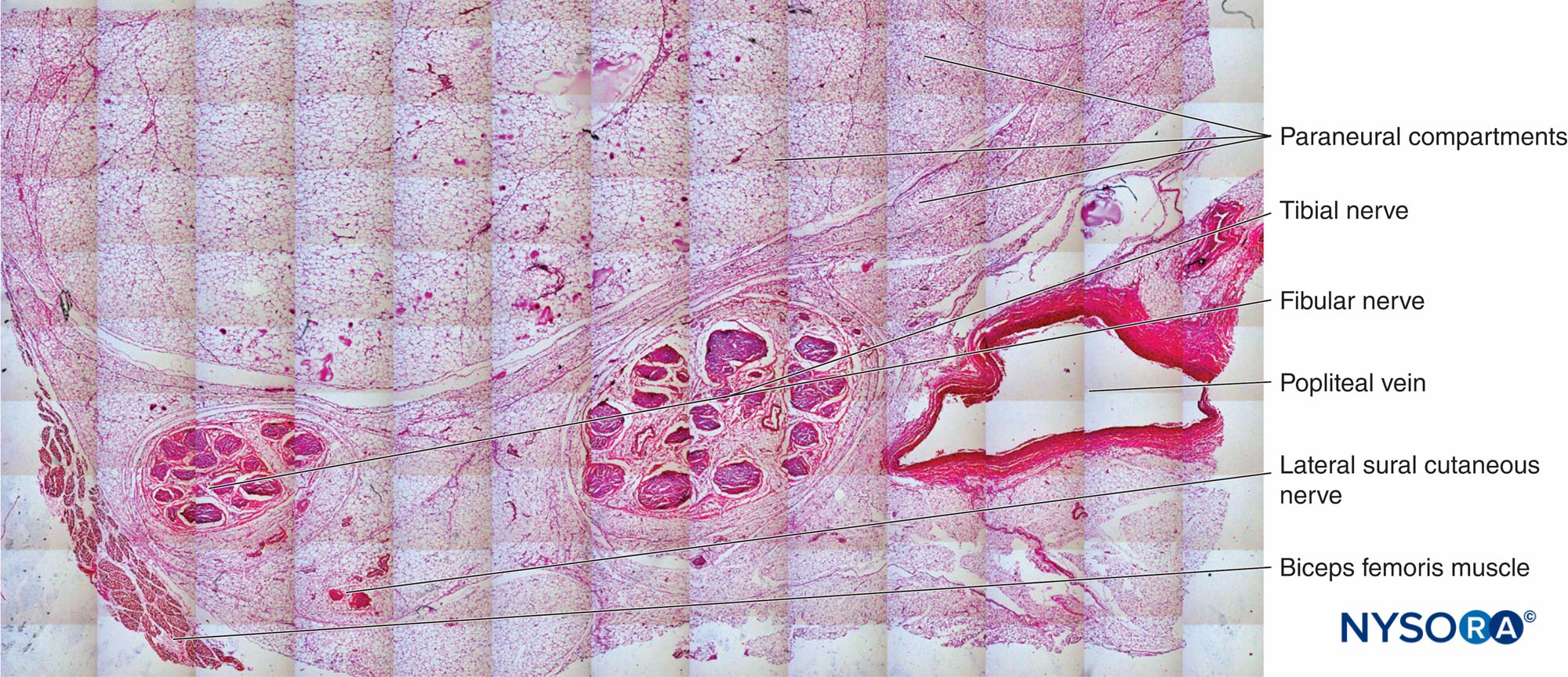
図25 膝窩の脛骨神経、腓骨神経およびその傍神経。 ヘマトキシリン-エオシン。 (Reina MAの許可を得て複製:局所麻酔および疼痛医学のための機能解剖学のアトラス。ニューヨーク:Springer; 2015年。)
臨床診療では、局所麻酔薬の超音波ガイド下注射により、この層と神経の間の空間が拡大して同心形状を示しているため、傍神経節の間接的な識別が可能になります。 筋束、筋束のグループ、神経、およびより複雑な神経構造を取り巻く神経層は同様の形態を持ち、それらは主にコラーゲン線維で構成されています。 したがって、各神経筋膜の解剖学的位置に基づく現在の宗派はかなり混乱しているように見えることに注意して、用語を統一することは合理的に思えるかもしれません。 ただし、現在の混乱を避けるために、神経上膜と傍神経上膜は、用語を使用するのが最善の場合があります。 神経上膜と傍神経膜はどちらも同様の機能を持っており、神経の絶縁と損傷からの保護が含まれます。 傍神経区画は、体の動きを制御する神経の縦方向の変位を促進します。 この動きは、その形状を変えることによって横方向の圧縮を中和するために必要です。 組織が外部刺激にさらされると、組織は反応し、束間線維症を引き起こします。
坐骨神経の解剖学的特徴に関連して、Andersen et alは、坐骨神経を取り巻く鞘が、肉眼的にも顕微鏡的にも神経上膜とは明らかに異なる薄い透明な構造であることを発見しました。 鞘は、神経に沿った注入物の広がりを促進しました。 しかし、その突起は神経を完全に取り囲んでいませんでした。 坐骨神経の周りの傍神経節の内層は、その鞘の同様の構造を持っていました。 Vloka et alは、一般的なエピネウラルシースという用語を使用しました。 トランらは、巨視的分裂に近い脛骨神経と腓骨神経が共有する総腓骨神経区画を満たす「分岐部」での「エピネウラル下」注射と呼ばれるものに関連して、坐骨神経ブロックの有効性を比較しました。
Orebaugh et alは、斜角筋間領域での針先の配置と麻酔液の注入が神経上膜内で頻繁に行われたことを報告しました。 これは、神経ブロックの約50%で発生し、筋束または軸索の損傷の形跡はなく、針がそれらを横切ったことを示唆する色素の痕跡も筋束内にありませんでした。 Spinner et alは、色素の上皮内注射が最も抵抗の少ない経路に沿ってその解剖をもたらすことを実証し、上神経区画の間に解剖学的制約が存在することを示唆している。 スピナーが内部神経上膜を注入したとき、染料は同じコンパートメント内で拡張しましたが、共通の外部神経上膜空間を越えたり、拡張したりしませんでした。 したがって、「神経内注射」の概念は、末梢神経間のかなりの解剖学的変動による単一神経の研究に基づく外挿を避けて、検査する神経ごとに改訂する必要があります。
NYSORAのヒント
プレクサスと末端部位の両方の末梢神経は、脂肪組織の同心クラスターに囲まれています。 これは、神経周囲注射が低い開口部注射圧をもたらす理由を説明しています。
末梢神経ブロック
麻酔薬の軸索への拡散は、結合組織鞘(例えば、神経周膜、ミエリン)の存在と特徴、および筋束内の軸索のサイズと位置に影響されます。 静脈内末梢麻酔(ビアブロック)中、局所麻酔薬は神経内毛細血管網を介して末梢神経終末に到達する可能性が最も高いです。 神経周膜および神経内毛細血管内皮は、内皮細胞間および神経周囲細胞間の密着結合のおかげで、軸索を異物から保護します。 神経の上膜の外側に注入された局所麻酔薬は、軸索に到達するために神経上膜と神経周膜の両方を通過する必要があります。 その後、注入された麻酔薬のごく一部のみが軸索と直接接触し、発症の遅延、不完全、または神経ブロックの失敗につながります。 たとえば、ポピッツと共同研究者は、ラットの坐骨神経に1%のリドカインを注射し、ブロックが完了すると、局所麻酔薬の神経内量が注射された用量の約1.6%になることを発見しました。
概要
末梢神経の結合組織の構成と配置は、末梢神経の保護と局所麻酔の実践において主要な役割を果たします。 結合組織の特性と変動性は、神経ブロック注射中の局所麻酔薬の広がりに実質的に影響を与える可能性があり、したがって神経ブロックのダイナミクスと品質に影響を与える可能性があります。 この主題に関連する補足ビデオはで見つけることができます 神経ブロックビデオの解剖学。
参考文献
- Reina MA、LópezA、Villanueva MC、DeAndrésJA、LeónGI:末梢神経、その鞘、およびそれらの血管新生の形態。 Rev Esp Anestesiol Reanim 2000; 47:464–475。
- Reina MA、Wikinski J、Prats-Galino A、MachésF:Morfologiadelnervioperiférico。 In:Wikinski J、Reina MA、Bollini C、et al(eds):Diagnóstico、prevenciónytratamiento delascomplicacionesneurológicasasociadasconlaanestesiaregionalperiféricaycentral。 Panamericana Ed、2011年、71〜86ページ。
- Reina MA、Arriazu R、Collier CB、Sala-Blanch X:神経ブロックの実践に臨床的に関連するヒト末梢神経の組織学および電子顕微鏡検査。 Rerv Esp Anestesiol Reanim 2013; 60:552–562。
- Reina MA、De Andres JA、HernándezJM、et al:くも膜下神経根から末梢神経への神経外構造の連続的な変化、麻酔ブロックへの影響、および急性術後疼痛の治療。 Eur J Pain Suppl 2011; 5:377–385。
- サンダーランドS、マーシャルRD、スワニーWE:サーカムフレックス筋皮神経および閉塞神経の神経内トポグラフィー。 Brain 1959; 82:116129–XNUMX。
- サンダーランドS、レイLJ:坐骨神経の神経内トポグラフィーと人間の膝窩部。 Brain 1948; 71:242–258。
- サンダーランドS:橈骨神経、正中神経、尺骨神経の神経内トポグラフィー。 Brain 1945; 68:243–255。
- サンダーランドS:Troncosnerviososperiféricos。 Salvat Ed、1985年、31〜60ページ。
- ボイドIA、デイビーMR:末梢神経の構成。 チャーチルリビングストーン、1968年。
- Friede RL、Bischhausen R:走査型電子顕微鏡で明らかになった末梢神経の神経内膜コラーゲンの組織化。 J Neurol Sci 1978; 38:83–89。
- Reina MA、LópezA、DeAndrésJA:末梢神経の血液脳関門。 Rev Esp Anestesiol Reanim 2003; 50:80–86。
- DeAndrésJA、Reina MA、LópezA、Sala-Blanch X、Prats A. Blocsnerveuxpériphériques、感覚異常および神経内注射。 Prat Anesth Reanim 2010; 14:213–221。
- Olsson Y、Kristensson K:神経への外傷後のタンパク質トレーサーへの拡散障壁としての神経周膜。 Acta Neuropath 1973; 23:105–111。
- サンダーランドS、ブラッドリーKC:末梢神経の神経周膜。 Anat Rec 1952; 113:125–142。
- Olsson Y、Resse TS:蛍光および電子顕微鏡で研究されたマウス坐骨神経における神経血管および神経周膜の透過性。 J Neuropathol Exp Neurol 1971; 30:105–119。
- Soderfeldt B、Olsson Y、Kristensson K:ヒト末梢神経のタンパク質トレーサーへの拡散障壁としての神経周膜。 Acta Neuropath 1973; 25:120–126。
- Llewelyn JG、Thomas PK:ストレプトゾトシン糖尿病ラットにおける神経周膜ナトリウム-カリウムATPアーゼ活性。 Exp Neurol 1987; 97:375–382。
- Lundborg G:外傷、浮腫の形成、神経機能に関連する神経内微小血管の構造と機能。 J Bone Joint Surg 1975; 57:938–948。
- サンダーランドS.神経周膜の破裂が封じ込められた神経線維に及ぼす影響。 Brain 1946; 69:149–152。
- サンダーランドS:末梢神経の結合組織。 Brain 1965; 88:841–854。
- サンダーランドS:末梢神経の脂肪組織。 Brain 1945; 68:118–122。
- Reina MA、LópezA、DeAndrésJA:末梢神経内の脂肪組織。 人間の坐骨神経の研究。 Rev Esp Anestesiol Reanim 2002; 49:397–402。
- Krstic R. Die Gewebe des Menschen undderSeaugetiere。 スプリンガー、1978年。
- Andersen HL、Andersen SL、Tranum-Jensen J.坐骨神経の傍神経鞘内への注射:超音波画像、肉眼解剖学、および組織学的分析の直接比較。 Reg AnesthPainMed。 2012; 37:410–414。
- Vloka JD、Hadzic A、Lesser JB、et al:膝窩の神経の一般的なエピネウラルシースと坐骨神経ブロックへの影響の可能性。 Anesth Analg 1997; 84:387–390; 膝窩における坐骨神経の分裂:膝窩神経ブロックの解剖学的意味。 Vloka JD、HadzićA、April E、Thys DM AnesthAnalg。 2001年92月;1(215):217-XNUMX。
- LangJ.神経の結合組織と血管について。 Z Anat Entwicklungsgesch 1962; 123:61–79。
- Van Beek A、Kleinert HE:実用的な神経障害。 Orthopedic Clin North Am 1977; 8:377–386。
- Sala-Blanch X、Reina MA、Ribalta T、Prats-Galino A.坐骨神経の構造と命名法:神経上膜から傍神経上膜。 これは新しいパラダイムですか? Reg Anesth Pain Med 2013; 38:463–465。
- 29. Sala-Blanch X、Vandepitte C、Laur J、et al:神経周囲注射と神経内注射の実際的なレビュー:標準的な命名法の必要性。 Int Anesth Clin 2011; 49:1–12。
- Millesi H、Hausner T、Schmidhammer R、Trattnig S、Tschabitscher M:末梢神経幹および筋束の受動的な運動性を提供する解剖学的構造。 Acta Neurochir Suppl 2007; 100:133–135。
- Reina MA(ed):局所麻酔および疼痛医学のための機能解剖学のアトラス:人間の構造、超構造および3D再構成画像。 Springer、2015年。
- Tran de QH、Dugani S、Pham K、Al-Shaafi A、Finlayson RJ:エピネウラル下と従来の超音波ガイド下膝窩坐骨神経ブロックのランダム化比較。 Reg Anesth Pain Med 2011; 36:548–552。
- Orebaugh SL、McFadden K、Skorupan H、Bigeleisen PE:超音波ガイド下の斜角筋間針先配置における上皮下注射。 Reg Anesth Pain Med 2010; 35:450–454。
- Spinner RJ、Wang H、Carmichael SW、Amrami KK、Scheithauer BW:上神経コンパートメントと神経内ガングリオン嚢胞伝播におけるそれらの役割:実験的研究。 Clin Anat 2007; 20:826–833。
- Ip V、Tsui B.円周方向の広がりではなく、傍神経鞘を介した注射は、安全で効果的な坐骨神経ブロックを促進します。 Reg Anesth Pain Med 2013; 38:373。
- Ip VH、ブリティッシュコロンビア州ツイ。 2つの石で1羽の鳥を殺す:膝窩坐骨神経ブロック中の分岐部への注射。 Reg Anesth Pain Med 2011; 36:633–634。
- Sala-Blanch X、LópezA、Prats-Galino A. Vloka坐骨神経鞘:先見の明のある人へのオマージュ。 Reg AnesthPainMed。 2015年40月-2月;174(XNUMX):XNUMX。
- Popitz-Bergez S、Lee-Soon S、Strichartz GR、Thalhammer JG:末梢神経ブロック中の機能障害と神経内局所麻酔薬との関係。 ラット坐骨神経の研究。 Anesthesiology 1995; 83:583–592。

